Dysregulated Inflammatory Cytokine Responses in Individuals with Colorectal Cancer: An Exploration of Immune Imbalance.
- Nikolay Shakhpazyan , Avtsyn Research Institute of Human Morphology, Petrovsky National Research Center of Surgery, 119435, Moscow, Russia;
- Nikolay Sadykhov , Research Institute of Human Morphology, Petrovsky National Research Center of Surgery, 119435, Moscow, Russia;
- Liudmila Mikhaleva , Avtsyn Research Institute of Human Morphology, Petrovsky National Research Center of Surgery, 119435, Moscow, Russia;
- Arkady Bedzhanyan , Department of Abdominal Surgery and Oncology II (Coloproctology and Uro-Gynecology), Petrovsky National Research Center of Surgery, 119435, Moscow, Russia;
- Alexandra Konyukova , Avtsyn Research Institute of Human Morphology, Petrovsky National Research Center of Surgery, 119435, Moscow, Russia;
- Ksenia Maslyonkina , Avtsyn Research Institute of Human Morphology, Petrovsky National Research Center of Surgery, 119435, Moscow, Russia;
- Andrey Kontorschikov , Avtsyn Research Institute of Human Morphology, Petrovsky National Research Center of Surgery, 119435, Moscow, Russia;
- Alexander Orekhov , Laboratory of Angiopathology, Institute of General Pathology and Pathophysiology, Moscow 125315, Russia; Institute for Atherosclerosis Research, Moscow 121096, Russia;
Article Information:
Abstract:
This study examines pro-inflammatory cytokines tumor necrosis factor alpha (TNF-α) and interleukin 1beta (IL-1β) in colorectal cancer (CRC). Twelve CRC patients and 9 control individuals came in [Name of the clinic] from April to June 2023 were enrolled. Study participants were recruited using the Convenience Sampling method. The CRC (colorectal cancer) patient group consisted of individuals with morphologically confirmed colorectal cancer who were scheduled for radical surgical treatment, prior to the administration of any pharmacological or radiation therapy. Control group individuals were volunteers of comparable age and sex, without oncological or chronic diseases affecting the immune system or related to inflammation. Monocytes were isolated and analyzed through Double lipopolysaccharide (LPS) Stimulation Assay and enzyme-linked immunosorbent assay (ELISA). Results indicate diminished TNF-α secretion post-initial LPS stimulation and no recovery upon re-stimulation in CRC monocytes (132 pg/ml in the control group versus 4 pg/ml in the CRC group, p=0.007). Conversely, elevated IL-1β levels were observed, particularly after re-stimulation (115 pg/ml in the control group versus 249 pg/ml in the CRC group, p=0.01). The findings suggest altered cytokine dynamics in CRC and potential avenues for targeted therapeutic interventions.
Keywords:
Article :
INTRODUCTION:
Colorectal cancer (CRC) stands as a significant worldwide health concern, occupying the position of the third most prevalent form of cancer. Central to deciphering the mechanisms underlying CRC is the complex interplay among the tumor milieu, immune response, and inflammatory processes [1]. Cytokines such as Tumor Necrosis Factor-alpha (TNF-α) and Interleukin-1beta (IL-1β) are central to this dynamic, each having distinct roles yet sometimes overlapping effects. TNF-α and IL-1β exhibit several significant differences in their mechanisms of secretion and activity. Both cytokines are regulated at the transcriptional level by NF-κB (Nuclear Factor kappa-light-chain-enhancer of activated B cells), a transcription factor that plays a key role in inflammatory responses [2]. TNF-α is synthesized as a transmembrane protein and is cleaved into its soluble form by the enzyme TNF-α converting enzyme (TACE) [3]. The production of TNF-α is rapidly induced in monocytes by various stimuli such as lipopolysaccharide (LPS), viruses, and other cytokines. TNF-α mRNA is constitutively present in monocytes, allowing for a swift response upon stimulation [4]. Unlike TNF-α, IL-1β is not stored pre-formed in cells. Its production involves two steps: transcription of the IL-1β gene into mRNA and then translation into a precursor protein, pro-IL-1β. The processing of pro-IL-1β into its active form requires the activation of the inflammasome, a multi-protein complex that activates caspase-1 [5]. This activation is typically a response to a range of stimuli including microbial products, ATP, or crystalline substances. The cytokines also differ in their mechanisms of release from the cell. The soluble form of TNF-α is released into the extracellular space after cleavage [6]. The release of IL-1β is not as straightforward as that of TNF-α. Since IL-1β lacks a signal peptide for classical secretion, it is released through mechanisms such as exocytosis of secretory lysosomes, shedding of microvesicles, or pyroptosis (a form of cell death associated with inflammation) [7, 8]. Both cytokines play critical roles in inflammation and immune response. However, their functions, while overlapping in some aspects, are distinct in their pathways and effects. TNF-α is recognized for its multifaceted and significant role in the carcinogenesis of colorectal cancer. Research indicates that TNF-α can exhibit pro-tumorigenic properties. For instance, colorectal cancer cells under the influence of TNF-α in culture tend to acquire a pro-metastatic phenotype, demonstrating enhanced proliferation and survival. This involves various mechanisms, including the activation of tumor cells through the STAT3 signaling pathway, leading to increased proliferative activity and survival [9]. The pro-tumorigenic effects of extracellular vesicles containing TNF-α are attributed to their impact on cellular processes involving NF-kB, LAMB3, PI3K/AKT signaling pathways [10]. The regulation of microRNA miR-21 expression is identified as crucial [11]. A pro-tumorigenic effect, marked by increased cell migration and invasion, is observed through the activation of TROP-2 (tumor-associated calcium signal transducer protein-2) expression by this cytokine [12]. Conversely, the anti-tumorigenic effects of anti-TNF-α antibodies have been observed in an orthotopic mouse model of colorectal cancer. This includes both a direct anti-tumorigenic effect and an influence on the stromal microenvironment, including reduced angiogenesis [13]. TNF-α plays a significant role in the development of colitis-associated colorectal cancer [14]. It has been noted that the level of TNF-α positively correlates with the presence of colorectal cancer metastases in lymph nodes in patients with tumor recurrence [15]. Additionally, the level of TNF-α is associated with the biological properties of the tumor, and elevated levels in the blood are observed in low-differentiated colorectal cancer [16]. However, the role of TNF-α in carcinogenesis is complex, and a reduction in TNF-α levels can also have a pro-tumorigenic effect [17]. Specifically, a decrease in TNF-α production by tumor-recruited monocytes leads to tumor progression related to myeloid to endothelial differentiation [18]. The influence of TNF-α levels on tumor pathogenesis may be linked to features of the tumor microenvironment. One mechanism influencing the anti-tumorigenic immune response is the polarization of tumor-associated macrophages (TAMs). It is known that M1 polarization of TAMs promotes the development of innate and adaptive immune responses against tumors, whereas M2 polarization supports tumor growth [19-21]. M1 polarization is characterized by the secretion of pro-inflammatory cytokines, including TNF-α, by TAMs [22, 23]. A decrease in TNF-α secretion may indirectly indicate a tendency towards M2 polarization of TAMs. Considering the migration of monocytes from blood to the tumor site and their subsequent transformation into TAMs, it is of interest to study the characteristics of the inflammatory response of monocytes in cancer patients. IL-1β exhibits a complex and, at times, contradictory influence on tumor biology. A well-established pro-tumoral mechanism of this cytokine involves its impact on the tumor stroma, encompassing remodeling and angiogenesis [24, 25]. Recent studies have revealed contradictory effects of IL-1β on the polarization of TAMs. Notably, IL-1β, a pro-inflammatory cytokine, has been shown to shift the balance towards M2 polarization of macrophages through the activation of the STAT3 signaling pathway via chemokine (C-X-C motif) ligand 8 (CXCL8) [26]. Additionally, IL-1β is capable of inducing apoptosis in M1-polarized macrophages, thereby potentially favoring a shift towards M2 polarization, which may have a pro-tumoral effect [27]. On the other hand, it has been observed that inhibition of IL-1β secretion, triggered by low expression of SHP-2, is associated with M2 polarization of TAMs in colorectal tumors [28]. The complexity of the tumor microenvironment and the multifaceted roles of cytokines such as IL-1β underscore the necessity for further research to attain a comprehensive understanding of their functions and interactions within the context of cancer biology. This study aims to scrutinize the roles of TNF-α and IL-1β, particularly their secretion by blood monocytes, to elucidate their individual and collective contributions to CRC.
Materials and Methods:
Participant Recruitment and Group Formation
This is an observational study were observed and recorded the differences in cytokine dynamics between the CRC group and the control group. Participants in this study who visited [Name of the Clinic] from April to June 2023 were enrolled. The study participants were recruited using the Convenience Sampling method. Convenience sampling is a non-probabilistic sampling method where participants are selected based on their availability and willingness to take part in the study, rather than being randomly chosen. Convenience sampling is chosen for this study as it efficiently targets a specific patient group, is resource-effective for short-communication research, and ethically aligns with recruiting willing participants with specific medical conditions. Common inclusion criteria for all participants included being older than 18 and having a conscious desire to participate in the study, as expressed in a signed informed consent. Common exclusion criteria for all participants were critical cardiovascular conditions: Stage III-IV tissue ischemia, acute cerebral circulatory disturbance, acute coronary syndrome, myocardial infarction, and chronic heart failure of class III and IV according to The New York Heart Association (NYHA); Critical clinical conditions not directly related to cardiovascular diseases, including the need for urgent surgeries, chronic renal failure of stages IV-V (creatinine clearance < 30 ml/min according to the Cockroft-Gault formula); Severe disability of the patient (4 or higher on the modified Rankin Scale); Systemic autoimmune diseases in the anamnesis, including: rheumatoid arthritis, systemic lupus erythematosus, autoimmune thyroiditis, autoimmune vasculitides, ulcerative colitis. There were formed two groups: CRC (colorectal cancer) patient group and Control group The CRC (colorectal cancer) patient group consisted of individuals with morphologically confirmed colorectal cancer who were scheduled for radical surgical treatment, prior to the administration of any pharmacological or radiation therapy. The stage of the disease was confirmed by examining the surgical material. The Control group comprised volunteers, without oncological or chronic diseases affecting the immune system or related to inflammation, and without significant weight loss (more than 10%) over the past year. We used demographic matching approach - the Control group participants were matched for age and gender with the patient group.
Study Participants
The investigation included a cohort of 21 subjects, comprising 12 individuals recently diagnosed with colorectal adenocarcinoma and an additional 9 controls who were matched for age and gender. Exclusion criteria filtered out individuals with specific acute and chronic diseases. Prior to any therapeutic intervention. All procedures, including comprehensive pathomorphological analysis post-surgery, adhered to the 1975 Helsinki Declaration and its 2013 update. Written informed consent was obtained from all participants.
Monocyte Culture Isolation and Double LPS Stimulation Assay
Mononuclear cells were extracted from blood samples of 30 ml utilizing Ficoll density gradient centrifugation, succeeded by the immunomagnetic separation of CD14+ cells employing LS Columns and CD14+ MicroBeads (Miltenyi Biotec Inc., USA). The purity of isolated monocytes confirmed by flow-cytometry (97.2±1.82% CD14+ cells) (Fig. 1). The isolated CD14+ monocytes were subsequently cultured in a concentration of one million cells per ml utilizing X-VIVO serum-free medium (Lonza, Switzerland), supplemented with gentamicin, L-glutamine, and phenol red, under conditions of 37°C and a 5% CO₂ atmosphere. Cultivation performed in cultural plates with 0.5 ml volume of cell-media suspension (500 000 cells) per a well. Triplicate cultures were established per patient. LPS stimulation was conducted at 1 µg/ml concentration of LPS. Two wells received LPS on day 1, with the third serving as a control. Cytokine levels were assessed at 24-hour intervals and after a medium refresh. Re-stimulation with LPS occurred in one well on day 6. Medium was collected on day 7 for final cytokine analysis and stored at -70°C until assayed for TNF-alpha and IL-1beta concentrations.
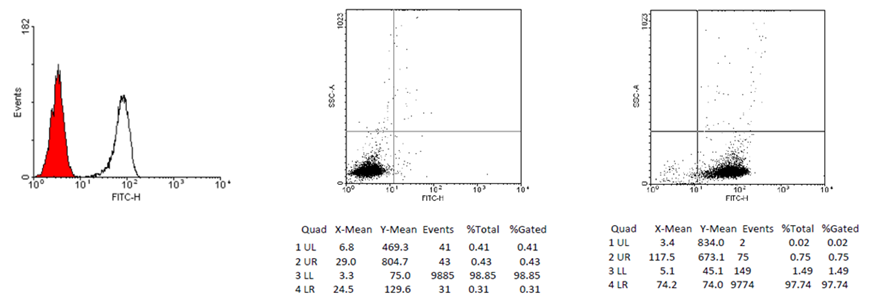
Figure 1: Flow-cytometry analysis of a sample immunomagnetic separated
cells, 97.74% cells are CD14+.
Measurement of TNF-alpha and IL-1beta Concentrations
Concentrations of TNF-alpha and IL-1beta in the culture medium were ascertained utilizing ELISA kits (Human TNF-alpha/TNFSF1A DuoSet ELISA and Human IL-1beta/IL-1F2 Duo-Set ELISA; R&D Systems Inc., USA). The evaluations were structured to encompass four distinct scenarios: 1) endogenous cytokine concentrations after a 24-hour period in the absence of LPS (termed non-stimulated), 2) cytokine concentrations at the 24-hour mark subsequent to initial LPS exposure (designated as first stimulation), 3) baseline cytokine concentrations on day 7 in cultures that had not received an additional LPS stimulation (termed non-re-stimulated), and 4) cytokine concentrations on the seventh day following a 24-hour period of LPS re-stimulation (designated as re-stimulation).
Statistical Analysis
Statistical evaluations were executed utilizing SPSS Statistics software, version 26.0. The distribution of data was appraised via the Shapiro-Wilk test to choose parametric or non-parametric statistical tests. Quantitative outcomes are delineated as median values accompanied by interquartile ranges. Disparities between the colorectal adenocarcinoma cohort and the control group were scrutinized employing the Mann-Whitney U-test. Additionally, bootstrap analysis [29] was conducted to estimate the variability and confidence intervals of our key statistics. This involved resampling our data with replacement to create numerous bootstrap samples, each equal in size to the original dataset. The resampling process was iterated 10,000 times, allowing for the robust estimation of the variability and confidence intervals for the median cytokine levels. A predetermined alpha level of 0.05 was set for all statistical tests to determine significance.
Results:
Participant group characteristics are presented in Table 1.
In our study, we aimed to investigate the differential cytokine responses of blood monocytes in CRC patients and healthy controls to LPS stimulation. We focused on TNF-alpha and IL-1beta, given their pivotal roles in modulating tumor inflammation and their distinct cellular mechanisms [30-32]. Upon LPS stimulation, we found that monocytes from CRC patients exhibited an initial secretion of TNF-α, followed by an exhaustion in its secretion. Specifically, a lower basal level was observed after a 7-day culture and minimal increase upon LPS re-stimulation (Table 2). Conversely, IL-1beta secretion showed elevated baseline levels and increased responsiveness to re-stimulation in CRC patients (Table 2). These findings raise questions about altered immune responses in CRC. Monocytes in CRC patients may undergo a form of "tolerization," reducing their efficacy against tumors and potentially fueling the tumor microenvironment. The abnormal cytokine secretion patterns observed underscore the complexity of CRC pathogenesis and the role of chronic inflammation therein.
Table 2. Quantification of TNF-α and IL-1β Release
|
Analyte |
Time point |
Condition |
Control |
CRC |
p-value (Mann-Whitney U test) |
|
TNF-α concentration (pg/ml) |
Concentration at 24 hours |
Non-stimulated with LPS |
111 (85-465) |
108 (63-210) |
0.743 |
|
|
Concentration at 24 hours |
Initial LPS Exposure |
4498 (2456-7446) |
3323 (2699-5708) |
0.842 |
|
|
Concentration at 7 days |
LPS Non-Stimulation Condition |
136 (79-179) |
4 (2-5) |
0.001* |
|
|
Concentration at 7 days |
Re-stimulation with LPS |
132 (79-182) |
7 (4-10) |
0.007* |
|
IL-1β concentration (pg/ml) |
Concentration at 24 hours |
LPS Non-Stimulation Condition |
96 (62-209) |
273 (259-333) |
0.005* |
|
|
Concentration at 24 hours |
Initial LPS Exposure |
1084 (422-4105) |
1313 (1084-1626) |
0.328 |
|
|
Concentration at 7 days |
LPS Non-Stimulation Condition |
115 (87-162) |
188 (176-212) |
0.113 |
|
|
Concentration at 7 days |
Re-stimulation with LPS |
115 (82-180) |
249 (197-328) |
0.01* |
LPS, lipopolysaccharide; CRC, colorectal cancer. Statistical significance is indicated by an asterisk.
DISCUSSION :
Our study elucidates differential cytokine secretion patterns in blood monocytes from CRC patients as compared to healthy controls. Specifically, monocytes in CRC show an exhaustion in TNF-α secretion and an elevated IL-1β secretion following LPS stimulation. As we mentioned, these cytokines serve distinct roles in inflammatory processes, regulated via unique pathways and kinetic profiles [19-23, 27, 28, 30-32]. Specifically, we noted exhaustion in TNF-α secretion and elevated IL-1β secretion following LPS stimulation. In CRC patients, the monocytes showed a reduced ability to secrete TNF-α after being stimulated with LPS. TNF-α is a cytokine involved in systemic inflammation and is one of the cytokines that make up the acute phase reaction. The term "exhaustion" here implies that these cells are less capable of producing TNF-α after stimulation, which could be significant in understanding the immune response in CRC. Conversely, these monocytes exhibited an increased production of IL-1β following LPS stimulation. IL-1β is another important cytokine in the body's inflammatory response, playing a role in cell proliferation, differentiation, and apoptosis (programmed cell death). These cytokines (TNF-α and IL-1β) have different roles in the body's inflammatory processes and are regulated through distinct pathways and kinetic profiles. This means that they are activated through different mechanisms in the body and have different timings in terms of their production and action [4, 5]. As an example, the TNF-α pathway, primarily modulated through NF-κB signaling, is involved in acute inflammation and cellular survival mechanisms, including apoptosis via Fas receptors [33]. IL-1β, conversely, activates immune cells and is regulated through the NLRP3 inflammasome pathway [34-35]. The secretion kinetics of these cytokines are divergent as well, with TNF-α peaking within 1-2 hours after LPS exposure, whereas IL-1β reaches a peak at 4-6 hours [33-35]. The altered secretion profiles in CRC potentially align with the theory that CRC is an inflammation-associated malignancy. This leads us to several hypotheses that warrant further investigation:
1. Acquired immune tolerance in monocytes could serve as a protective mechanism for tumor cells against immune attack [36]. This could particularly be relevant given that IL-1β secretion is elevated in CRC patients, necessitating a broader cytokine panel for a complete understanding. 2. Abnormalities in key signaling pathways such as NF-κB and MAPK in monocytes could be pivotal in understanding the observed phenotypes [37]. 3. The tumor microenvironment, a dynamic entity involving various soluble factors and exosomes, could influence the reduced TNF-α re-secretion [38]. 4. A depletion in monocyte count in CRC patients could also contribute to the observed exhaustion of TNF-α secretion [39]. 5. Lastly, CRC-associated alterations in gut microbiota and intestinal permeability could be influencing monocyte interactions and subsequent cytokine secretion [39].
Collectively, these findings and hypotheses open avenues for future research aimed at elucidating the complex interplay between inflammation, immune responses, and CRC pathogenesis.
CONCLUSION :
In summary, the observed modulation in cytokine response among individuals with colorectal carcinoma implicates a contributory role in shaping the tumor milieu and related inflammatory mechanisms integral to CRC advancement. This aberrant cytokine profile serves as a prospective target for therapeutic interventions aimed at modulating the immune responsiveness in colorectal carcinoma patients.
6. Declarations
Conflict of interest
The authors have no conflicts of interest to declare.
Funding
This work was financially supported by the Russian Science Foundation (Grant number 25-25-00358).
BIBLIOGRAPHY:
1. Muthusami S, et al. (2021) Role of Inflammation in the Development of Colorectal Cancer. Endocr Metab Immune Disord Drug Targets 21:77-90. https://doi.org/10.2174/1871530320666200909092908
2. Liu, T., Zhang, L., Joo, D., & Sun, S. C. (2017). NF-κB signaling in inflammation. Signal transduction and targeted therapy, 2, 17023–. https://doi.org/10.1038/sigtrans.2017.23
3. Deora, A., Hegde, S., Lee, J., Choi, C. H., Chang, Q., Lee, C., Eaton, L., Tang, H., Wang, D., Lee, D., Michalak, M., Tomlinson, M., Tao, Q., Gaur, N., Harvey, B., McLoughlin, S., Labkovsky, B., & Ghayur, T. (2017). Transmembrane TNF-dependent uptake of anti-TNF antibodies. mAbs, 9(4), 680–695. https://doi.org/10.1080/19420862.2017.1304869
4. Wolf, Y., Shemer, A., Polonsky, M., Gross, M., Mildner, A., Yona, S., David, E., Kim, K. W., Goldmann, T., Amit, I., Heikenwalder, M., Nedospasov, S., Prinz, M., Friedman, N., & Jung, S. (2017). Autonomous TNF is critical for in vivo monocyte survival in steady state and inflammation. The Journal of experimental medicine, 214(4), 905–917. https://doi.org/10.1084/jem.20160499
5. Cardona Gloria, Y., & Weber, A. N. R. (2023). Inflammasome Activation in Human Macrophages: IL-1β Cleavage Detection by Fully Automated Capillary-Based Immunoassay. Methods in molecular biology (Clifton, N.J.), 2696, 239–256. https://doi.org/10.1007/978-1-0716-3350-2_16
6. Josephs, S. F., Ichim, T. E., Prince, S. M., Kesari, S., Marincola, F. M., Escobedo, A. R., & Jafri, A. (2018). Unleashing endogenous TNF-alpha as a cancer immunotherapeutic. Journal of translational medicine, 16(1), 242. https://doi.org/10.1186/s12967-018-1611-7
7. Li, Y., & Jiang, Q. (2023). Uncoupled pyroptosis and IL-1β secretion downstream of inflammasome signaling. Frontiers in immunology, 14,
1.1128358. https://doi.org/10.3389/fimmu.2023.1128358
8. Caielli, S., Balasubramanian, P., Rodriguez-Alcazar, J., Balaji, U., Wan, Z., Baisch, J., Smitherman, C., Walters, L., Sparagana, P., Nehar-Belaid, D., Marches, R., Nassi, L., Stewart, K., Fuller, J., Banchereau, J. F., Gu, J., Wright, T., & Pascual, V. (2023). An unconventional mechanism of IL-1β secretion that requires Type I IFN in lupus monocytes. bioRxiv: the preprint server for biology, 2023.08.03.551696. https://doi.org/10.1101/2023.08.03.551696
9. Wei W, Wang J, Huang P, Gou S, Yu D, Zong L. Tumor necrosis factor-α induces proliferation and reduces apoptosis of colorectal cancer cells through STAT3 activation. Immunogenetics. 2023 Apr;75(2):161-169. doi:
10. 1007/s00251-023-01302-y. Epub 2023 Mar 18. PMID: 36933092.
11. Xie BW, Guan B, Chen W, Zhou M, Gu Q, Liu Y, Yan D. Tumor-derived extracellular vesicles delivering TNF-α promotes colorectal cancer metastasis via the NF-kB/LAMB3/AKT axis by targeting SNAP23. Arch Biochem Biophys. 2023 Jun;741:109605. doi: 10.1016/j.abb.2023.109605. Epub 2023 Apr 21. PMID: 37086961.
12. Alotaibi AG, Li JV, Gooderham NJ. Tumour Necrosis Factor-Alpha (TNF-α)-Induced Metastatic Phenotype in Colorectal Cancer Epithelial Cells: Mechanistic Support for the Role of MicroRNA-21. Cancers (Basel). 2023 Jan 19;15(3):627. doi: 10.3390/cancers15030627. PMID: 36765584; PMCID: PMC9913347.
13. Zhao P, Zhang Z. TNF-α promotes colon cancer cell migration and invasion by upregulating TROP-2. Oncol Lett. 2018 Mar;15(3):3820-3827. doi:
1. 10. 3892/ol.2018.7735. Epub 2018 Jan 5. PMID: 29467899; PMCID: PMC5796310.
14. Takasago T, Hayashi R, Ueno Y, Ariyoshi M, Onishi K, Yamashita K, Hiyama Y, Takigawa H, Yuge R, Urabe Y, Oka S, Kitadai Y, Tanaka S. Anti-tumor necrosis factor-alpha monoclonal antibody suppresses colorectal cancer growth in an orthotopic transplant mouse model. PLoS One. 2023 Mar 30;18(3):e0283822. doi: 10.1371/journal.pone.0283822. PMID: 36996146; PMCID: PMC10062630.
15. Ba H, Jiang R, Zhang M, Yin B, Wang J, Li Z, Li B, Zhou X. Suppression of Transmembrane Tumor Necrosis Factor Alpha Processing by a Specific Antibody Protects Against Colitis-Associated Cancer. Front Immunol. 2021 Oct 5;12:687874. doi: 10.3389/fimmu.2021.687874. PMID: 34675913; PMCID: PMC8524043.
16. Grimm M, Lazariotou M, Kircher S, Höfelmayr A, Germer CT, von Rahden BH, Waaga-Gasser AM, Gasser M. Tumor necrosis factor-α is associated with positive lymph node status in patients with recurrence of colorectal cancer--indications for anti-TNF-α agents in cancer treatment. Anal Cell Pathol (Amst). 2010;33(3):151-63. doi: 10.3233/ACP-CLO-2010-0539. Erratum in: Cell Oncol (Dordr). 2011 Aug;34(4):407-9. PMID: 20978325; PMCID: PMC4605536.
17. Zheng J, Wang X, Yu J, Zhan Z, Guo Z. IL-6, TNF-α and IL-12p70 levels in patients with colorectal cancer and their predictive value in anti-vascular therapy. Front Oncol. 2022 Sep 26;12:997665. doi:
2. 10. 3389/fonc.2022.997665. PMID: 36226059; PMCID: PMC9549173.
18. Cruceriu D, et al. (2020) The dual role of tumor necrosis factor-alpha (TNF-α) in breast cancer: molecular insights and therapeutic approaches. Cell Oncol (Dordr) 43:1-18. https://doi.org/10.1007/s13402-019-00489-1
19. Li, B., Vincent, A., Cates, J., Brantley-Sieders, D. M., Polk, D. B., & Young, P. P. (2009). Low levels of tumor necrosis factor alpha increase tumor growth by inducing an endothelial phenotype of monocytes recruited to the tumor site. Cancer research, 69(1), 338–348. https://doi.org/10.1158/0008-
4.5472. CAN-08-1565
20. Basak, U., Sarkar, T., Mukherjee, S., Chakraborty, S., Dutta, A., Dutta, S., Nayak, D., Kaushik, S., Das, T., & Sa, G. (2023). Tumor-associated macrophages: an effective player of the tumor microenvironment. Frontiers in immunology, 14, 1295257. https://doi.org/10.3389/fimmu.2023.1295257
21. Zhong, X., Chen, B., & Yang, Z. (2018). The Role of Tumor-Associated Macrophages in Colorectal Carcinoma Progression. Cellular physiology and biochemistry: international journal of experimental cellular physiology, biochemistry, and pharmacology, 45(1), 356–365. https://doi.org/10.1159/000486816
22. Feng R, et al. (2018) Nrf2 activation drive macrophages polarization and cancer cell epithelial-mesenchymal transition during interaction. Cell Commun Signal 16:54. https://doi.org/10.1186/s12964-018-0262-x
23. Chong, L., Jiang, Y. W., Wang, D., Chang, P., Xu, K., & Li, J. (2023). Targeting and repolarizing M2-like tumor-associated macrophage-mediated MR imaging and tumor immunotherapy by biomimetic nanoparticles. Journal of nanobiotechnology, 21(1), 401. https://doi.org/10.1186/s12951-023-02122-8
24. Li, Y., Chen, Z., Han, J., Ma, X., Zheng, X., & Chen, J. (2022). Functional and Therapeutic Significance of Tumor-Associated Macrophages in Colorectal Cancer. Frontiers in oncology, 12, 781233. https://doi.org/10.3389/fonc.2022.781233
25. Kim, H. G., Huot, J. R., Pin, F., Belcher, D. J., Bonetto, A., & Nader, G. A. (2022). Metastatic or xenograft colorectal cancer models induce divergent anabolic deficits and expression of pro-inflammatory effectors of muscle wasting in a tumor-type-dependent manner. Journal of applied physiology (Bethesda, Md.: 1985), 133(6), 1273–1283. https://doi.org/10.1152/japplphysiol.00247.2022
26. Sharma, B. R., Karki, R., Sundaram, B., Wang, Y., Vogel, P., & Kanneganti, T. D. (2022). The Transcription Factor IRF9 Promotes Colorectal Cancer via Modulating the IL-6/STAT3 Signaling Axis. Cancers, 14(4), 919. https://doi.org/10.3390/cancers14040919
27. Shao, Y., Lan, Y., Chai, X., Gao, S., Zheng, J., Huang, R., Shi, Y., Xiang, Y., Guo, H., Xi, Y., Yang, L., & Yang, T. (2023). CXCL8 induces M2 macrophage polarization and inhibits CD8+ T cell infiltration to generate an immunosuppressive microenvironment in colorectal cancer. FASEB journal: official publication of the Federation of American Societies for Experimental Biology, 37(10), e23173. https://doi.org/10.1096/fj.202201982RRR
28. Zeng, Y. X., Chou, K. Y., Hwang, J. J., & Wang, H. S. (2023). The effects of IL-1β stimulated human umbilical cord mesenchymal stem cells on polarization and apoptosis of macrophages in rheumatoid arthritis. Scientific reports, 13(1), 10612. https://doi.org/10.1038/s41598-023-37741-6
29. Li, Z., Xi, J., Li, B., Liu, Y., Wang, G., Yu, B., Ma, H., Li, Z., & Zhang, Z. (2023). SHP-2-induced M2 polarization of tumor associated macrophages via IL-4 regulate colorectal cancer progression. Frontiers in oncology, 13,
3. 1027575. https://doi.org/10.3389/fonc.2023.1027575
30. Johnston, M. G., & Faulkner, C. (2021). A bootstrap approach is a superior statistical method for the comparison of non-normal data with differing variances. The New phytologist, 230(1), 23–26. https://doi.org/10.1111/nph.17159
31. Yao D, et al. (2019) Inflammation and Inflammatory Cytokine Contribute to the Initiation and Development of Ulcerative Colitis and Its Associated Cancer. Inflamm Bowel Dis 25:1595-1602. https://doi.org/10.1093/ibd/izz149
32. Kelley N, et al. (2019) The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int J Mol Sci 20:3328. https://doi.org/10.3390/ijms20133328
33. Chen L, et al. (2017) Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 9:7204-7218. https://doi.org/10.18632/oncotarget.23208
34. Liu T, et al. (2017) NF-κB signaling in inflammation. Signal Transduct Target Ther 2:17023. https://doi.org/10.1038/sigtrans.2017.23
35. Shalapour S, Karin M (2019) Pas de Deux: Control of Anti-tumor Immunity by Cancer-Associated Inflammation. Immunity 51:15-26. https://doi.org/10.1016/j.immuni.2019.06.021
36. Murray PJ (2018) Immune regulation by monocytes. Semin Immunol 35:12-
6.18. https://doi.org/10.1016/j.smim.2017.12.005
37. He X, et al. (2015) Baicalein attenuates inflammatory responses by suppressing TLR4 mediated NF-κB and MAPK signaling pathways in LPS-induced mastitis in mice. Int Immunopharmacol 28:470-6. https://doi.org/10.1016/j.intimp.2015.07.012
38. Liguori M, et al. (2011) Tumor-associated macrophages as incessant builders and destroyers of the cancer stroma. Cancers (Basel) 3:3740-61. https://doi.org/10.3390/cancers3043740
39. Schumak B, et al. (2015) Specific depletion of Ly6C(hi) inflammatory monocytes prevents immunopathology in experimental cerebral malaria. PLoS One 10:e0124080. https://doi.org/10.1371/journal.pone.0124080
40. Campbell C, et al. (2023) Crosstalk between Gut Microbiota and Host Immunity: Impact on Inflammation and Immunotherapy. Biomedicines 11:294. https://doi.org/10.3390/biomedicines11020294